Поперечнополосатая мышечная ткань. Поперечнополосатые мышцы
Мышечная ткань: виды, особенности строения, месторасположение в организме
Мышечные ткани (textus musculares) – это специализированные ткани, которые обеспечивают движение (перемещение в пространстве) организма в целом, а также его частей и внутренних органов. Сокращение мышечных клеток или волокон осуществляется с помощью миофиламентов и специальных органелл – миофибрилл и является результатом взаимодействия молекул сократительных белков.
Согласно морфункциональной классификации, мышечные ткани делят на две группы:
I – поперечнополосатая (исчерченная) мышечная ткань – содержит постоянно комплексы актиновых и миозиновых миофиламентов – миофибриллы и имеет поперечную исчерченность;
II – гладкая (неисчерченная) мышечная ткань – состоит из клеток, которые постоянно содержат только актиновые миофиламенты и не имеют поперечной исчерченности.
Поперечнополосатая мышечная ткань
Поперечнополосатая мышечная ткань подразделяется на скелетную и сердечную . Обе эти разновидности развиваются из мезодермы .
Поперечнополосатая скелетная мышечная ткань. Эта ткань образует скелетные мышцы, мышцы рта, глотки, частично пищевода, мышцы промежности и др. В разных отделах она имеет свои особенности. Обладает высокой скоростью сокращения и быстрой утомляемостью. Этот тип сократительной деятельности называется тетаническим . Поперечнополосатая скелетная мышечная ткань сокращается произвольно в ответ на импульсы, идущие от коры больших полушарий головного мозга. Однако часть мышц (межреберные, диафрагма и др.) имеет не только произвольный характер сокращения, но и сокращается без участия сознания под влиянием импульсов из дыхательного центра, а мышцы глотки и пищевода сокращаются непроизвольно.
Структурной единицей является поперечнополосатое мышечное волокно – симпласт, цилиндрической формы с округлыми или заостренными концами, которыми волокна прилежат друг к другу или вплетаются в соединительную ткань сухожилий и фасций.
Сократительным аппаратом их являются поперечнополосатые миофибриллы , которые образуют пучок волоконец. Это белковые нити, расположенные вдоль волокна. Длина их совпадает с длиной мышечного волокна. Миофибриллы состоят из темных и светлых участков – дисков . Так как темные и светлые диски всех миофибрилл одного мышечного волокна располагаются на одном уровне, образуется поперечная исчерченность; поэтому мышечное волокно называется поперечнополосатым.Темные диски в поляризованном свете имеют двойное лучепреломление и называются анизотропными, или А-дисками; светлые диски не имеют двойного лучепреломления и называются изотропными, или I-дисками.
Разная светопреломляющая способность дисков обусловлена их различным строением. Светлые (I) диски однородны по составу: образованы только параллельно лежащими тонкими нитями – актиновыми миофиламентами , состоящими преимущественно из белка актина , а также тропонина и тропомиозина . Темные (А) диски неоднородны: образованы как толстыми миозиновыми миофиламентами , состоящими из белка миозина , так и частично проникающими между ними тонкими актиновыми миофиламентами .
В середине каждого I–диска проходит темная линия, которая называется Z–линией, или телофрагмой . К ней прикрепляется один конец актиновых нитей. Участок миофибриллы между двумя телофрагмами называется саркомером . Саркомер – структурно-функциональная единица миофибриллы. В центре A-диска можно выделить светлую полосу, или зону Н , содержащую только толстые нити. В середине ее выделяется тонкая темная линия М, или мезофрагма . Таким образом, каждый саркомер содержит один А-диск и две половины I-диска .
Поперечнополосатая сердечная мышечная ткань. Образует миокард сердца. Содержит, как и скелетная, миофибриллы, состоящие из темных и светлых дисков. Состоит из клеток – кардиомиоцитов , связанных между собой вставочными дисками. При этом образуются цепочки кардиомиоцитов – функциональные мышечные волокна, которые анастомозируют между собой (переходят одно в другое), образуя сеть. Такая система соединений обеспечивает сокращение миокарда как единого целого. Сокращение сердечной мышцы непроизвольное , регулируется вегетативной нервной системой.
Среди кардиомиоцитов различают:
· сократительные (рабочие) кардиомиоциты – содержат меньше миофибрилл, чем скелетные мышечные волокна, но очень много митохондрий, поэтому сокращаются с меньшей силой, но долго не утомляются; с помощью вставочных дисков осуществляют механическую и электрическую связь кардиомиоцитов;
· атипичные (проводящие) кардиомиоциты – образуют проводящую систему сердца для формирования и проведения импульсов к сократительным кардиомиоцитам;
· секреторные кардиомиоциты – располагаются в предсердиях, способны вырабатывать гормоноподобный пептид – натрий-уретический фактор , снижающий артериальное давление.
Гладкая мышечная ткань
Развивается из мезенхимы, располагается в стенке трубчатых органов (кишечник, мочеточник, мочевой пузырь, кровеносные сосуды), а также радужке и цилиарном (ресничном) теле глаза и мышцах, поднимающих волосы в коже.
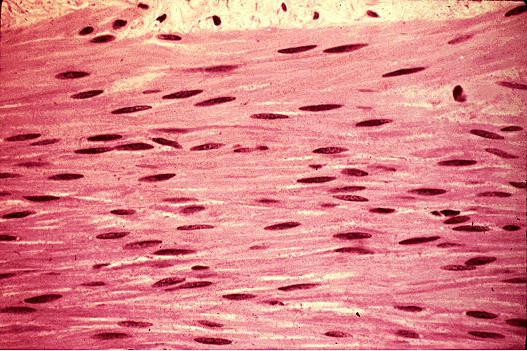
Гладкая мышечная ткань имеет клеточное строение (гладкий миоцит) и обладает сократительным аппаратом в виде гладких миофибрилл . Она сокращается медленно и способна длительно находиться в состоянии сокращения, потребляя относительно малое количество энергии и не утомляясь. Такой тип сократительной деятельности называется тоническим . К гладкой мышечной ткани подходят вегетативные нервы, и в отличие от скелетной мышечной ткани она не подчиняется сознанию, хотя и находится под контролем коры больших полушарий головного мозга.
Гладкомышечная клетка имеет веретенообразную форму и заостренные концы. В ней есть ядро, цитоплазма (саркоплазма), органеллы и оболочка (сарколемма). Сократительные миофибриллы располагаются по периферии клеток вдоль ее оси. Эти клетки плотно прилежат друг к другу. Опорным аппаратом в гладкой мышечной ткани являются тонкие коллагеновые и эластические волокна, расположенные вокруг клеток и связывающие их между собой.
Мышечная ткань (textus muscuIaris) представляет собой группу тканей (поперечнополосатая, гладкая, сердечная), имею-щих различное происхождение и строение, объединенных по функциональному признаку - способности сокращаться -- укорачиваться. Наряду с упомянутыми разновидностями мы-шечной ткани, образующейся из мезодермы (мезенхимы), в ор-ганизме человека выделяют мышечную ткань эктодермального происхождения - миоциты радужки глаза.
Исчерченная (поперечно-полосатая, скелетная) мышечная ткань образована цилиндрическими мышечными волокнами длиной от 1 мм до 4 см и более и толщиной до 0,1 мм. Каждое волокно представляет собой комплекс, состоящий из миосим-пласта и миосателлитоцитов, покрытых плазматической мем-браной, которую называют сарколеммой (от греч. sаг-kos - мясо). Снаружи к сарколемме прилежит базаль-ная пластинка (мембрана), образованная тонкими коллагеновы-ми и ретикулярными волокнами. Миосимпласт, находящийся под сарколеммой мышечного волокна, получил название сар-коплазмы. Он состоит из множества эллипсоидных ядер (до 100), миофибрилл и цитоплазмы. Удлиненные ядра, ориен-тированные вдоль мышечного волокна, лежат под сарколеммой. В саркоплазме имеется большое количество элементов зерни-стой эндоплазматической сети. Примерно 2/З сухой массы мы-шечного волокна приходится на цилиндрические миофибрил-лы, проходящие продольно почти через всю саркоплазму. Между миофибриллами располагаются многочисленные мито-хондрии с хорошо развитыми кристами и гликоген.
В поперечнополосатом мышечном волокне хорошо развита саркотубулярная сеть, которая образована двумя компонентами: канальцами эндоплазматической сети, расположенными вдоль миофибрилл (L-система), и Т-канальцами (Т-трубочками), начинающимися в области впячивания сарколеммы. Т-трубочки проникают в глубь мышечного волокна и образуют поперечные трубочки вокруг каждой миофибриллы. Т-трубочки играют важную роль в быстром проведении по-тенциала действия к каждой миофибрилле. Возникший в сарко-лемме мышечного волокна под воздействием нервного импуль-са потенциал действия распространяется по Т-трубочкам, а от них на незернистую эндоплазматическую сеть, канальцы кото-рой располагаются возле Т-трубочек, а также между миофиб-риллами.
Основную часть саркоплазмы мышечного волокна составля-ют специальные органеллы - миофибриллы. Каждая миофибрилла состоит из правильно чередующихся участков -темных анизотропных дисков А и светлых изотропных дисков I. В середине каждого анизотропного диска А проходит светлая зона - полоска Н, в центре которой находится линия М, или мезофрагма. Через середину диска I проходит линия Z - так называемая телофрагма. Чередование темных и светлых дисков в соседних миофибриллах, располагающихся на одном уровне, на гистологическом препарате скелетной мышцы создает впе-чатление поперечной исчерченности. Каждый темный диск об-разован толстыми миозиновыми нитями диаметром 10-15 нм. Длина толстых нитей около 1,5 мкм. Основу этих нитей (фила-ментов) составляет высокомолекулярный белок миозин. Каж-дый светлый диск образован из тонких актиновых нитей диа-метром 5-8 нм и длиной около 1 мкм, состоящих из низкомо-лекулярного белка актина, а также низкомолекулярных белков тропомиозина и тропонина. Участок миофибриллы между двумя телофрагмами (Z-ли-ниями) называют саркомером. Он является функцио-нальной единицей миофибриллы. Длина саркомера около 2,5 мкм, в него входят темный диск А и половинки примыкаю-щих к нему с двух сторон светлых дисков I. Таким образом, тон-кие актиновые нити идут от Z-линии навстречу друг другу и входят в диск А, в промежутки между толстыми миозиновыми нитями. При сокращении мышцы актиновые и миозиновые нити скользят навстречу друг другу, при расслаблении - двига-ются в противоположные стороны.
Саркоплазма богата белком миоглобином, который, подоб-но гемоглобину, может связывать кислород. В зависимости от толщины волокон, содержания в них миоглобина и миофиб-рилл различают так называемые красные и белые поперечнопо-лосатые мышечные волокна. Красные мышечные волокна (тем-ные) богаты саркоплазмой, миоглобином и митохондриями, од-нако в них мало миофибрилл. Эти волокна медленно сокраща-ются и долго могут быть в сокращенном (рабочем) состоянии. Белые мышечные (светлые) волокна содержат мало саркоплазмы, миоглобина и митохондрий, но в них много миофибрилл. Эти волокна сокращаются быстрее красных, но быстро «устают». У человека мышцы содержат оба типа волокон. Сочетание мед-ленных (красных) и быстрых (белых) мышечных волокон обеспечивает мышцам быстроту реакции (сокращение) и длитель-ную работоспособность.
Миосателлитоциты располагаются непосредствен-но над сарколеммой, но под базальной пластинкой (мембра-ной). Это уплощенные клетки с богатым хроматином крупным ядром. Каждый миосателлитоцит имеет центросому и малое число органелл; спиральных органелл (миофибрилл) у них нет. Миосателлитоциты являются стволовыми (ростковыми) клетка-ми поперечнополосатой (скелетной) мышечной ткани, они спо-собны к синтезу ДНК и митотическому делению.
Мышечные клетки обычно называют мышечными волокнами, потому что они постоянно вытянуты в одном направлении. Сократимость у мышечных клеток достигла наибольшего развития: они состоят из сокращающихся элементов, называемых миофибриллами, расположенных вдоль оси длинной волокна и придающих ему продольную исчерченность. Это характерно для всех видов мышечной ткани. Миофибриллы некоторых типов мышечной ткани состоят из перемежающихся плотноуложенных темных и светлых полос. В результате волокно имеет поперечную исчерченность и называется поперечно-полосатым мышечным волокном.
Классификация мышечных тканей проводится на основании строения ткани (гистологически): по наличию или отсутствию поперечной исчерченности, и на основании механизма сокращения - произвольного (как в скелетной мышце) или непроизвольного (гладкая или сердечная мышцы), т.е. по физиологическому признаку. Плазматическая мембрана мышечного волокна называется сарколеммой, и в зависимости от типа мышечной ткани она имеет различную степень развития.
Цитоплазма клеток мышечной ткани называется саркоплазмой, во всех трех видах мышечной ткани цитоплазма состоит из сократительных белков - актина и миозина и богата митохондриями, содержащими ферменты для активного метаболизма и сократительных движений мышечного волокна. Скелетная и сердечная мышечные ткани изобилуют гладким эндоплазматическим ретикулумом (саркоплазматическим реткулумом).
Классификация мышечной ткани.
А) По гистологическому признаку:
Неисчерченная:
Гладкая мышечная ткань
Исчерченная:
Сердечная мышца
Б) По физиологическому признаку:
Непроизвольная:
Гладкая мышечная ткань
Сердечная мышца
Произвольная:
Поперечно-полосатая мышечная ткань
Гладко-мышечная ткань.
Из гладкой мышечной ткани построены стенки висцеральных органов, мышечные оболочки кровеносных сосудов (артерий, артериол, вен, кроме венул). Длина мышечных волокон их колеблется от 20 до 500 мкм, а ширина в месте расположения ядра составляет около 5 мкм, в направлении обоих концов волокно истончается и заостряется. Ядро на поперечном срезе часто не визуализируется. Если же оно видно, то форма его сферическая. На продольном срезе ядро имеет овальную форму: при сокращении волокна оно может стать штопорообразным.
Сарколемму гладкой мышечной ткани очень трудно выявить при помощи световой микроскопии. Гладкие мышечные волокна окружены каркасом ретикулиновых волокон, погруженных в ШИК-положительное основное вещество. Считается, что ретикулиновые волокна служат ограничением для мышечного волокна, удерживая их при сокращении. При исследовании под электронным микроскопом выявляется, что с наружной стороны сарколемма покрыта базальной пластинкой. Гладкомышечные клетки могут увеличиваться в количестве с помошью митозов (гиперплазия), а также в размере (гипертрофия). Обычно гладкомышечные волокна располагаются в несколько слоев, в которых широкая часть одного волокна примыкает к суженным, вытянутым краям других волокон.
Гладкая мускулатура иннервируется вегетативной нервной системой: как симпатической, так и парасимпатической ее частей (стенка кишечника). В других случаях случаях сокращения мышечных органов осуществляется исключительно симпатической системой (стенка артерии), или парасимпатической (сфинктер зрачка). Нервные окончания могут располагаться только на поверхностных клетках слоя волокон: импульс проходит через слой от клетки к клетке через щелевые контакты (нексусы), области сужения межклеточного пространства с низким электрическим сопротивлением. Однако, в некоторых случаях, иннервируется каждая гладкомышечная клетка (радужная оболочка глаза и семявыносящий проток). В цитоплазме гладкомышечных клеток с помощью электронной микроскопии можно увидеть митохондрии, аппарат Гольджи, гликоген, шероховатый эндоплазматический ретикулум и рибосомы; эти органеллы располагаются у полюсов ядра. Цитоплазма также содержит интенсивно окрашенные, сигарообразные элементы, содержащие альфа-актин, объединенные в систему постоянных цитоплазматических филаментов промежуточного диаметра (10 нм) между тонкими актиновыми филаментами (7 нм толщиной) и толстыми миозиновыми филаментами (17 нм тощиной), которые также встречаются здесь. Сарколемма видна в местах инвагинации в продольные ряды пузырьков (кавеолы), которые тесно связаны с трубочками саркоплазматического ретикулума. Они регулируют концентрацию катионов кальция в клетке и проводят сократительные импульсы внутрь клетки (подобно Т-трубочкам поперечнополосатой мышечной ткани). В цитоплазме также встречаются микротрубочки, состоящие из тубулина.
Поперечно-полосатая мышечная ткань.
Волокна поперечно-полосатых мышц представляют собой многоядерные гигантские клетки цилиндрической формы: их длина колеблется от нескольких милиметров до нескольких сантиметров. Их толщина составляет от 10 до 150 мкм. Сарколемма достаточно развита, вдоль нее раположены многочисленные ядра, неравномерно разбросанные в пределах волокна. Они имеют овальную форму на продольном срезе и округлую на поперечном. Волокна характеризуются как продольной, так и поперечной исчерченностью, которая выражена значительнее. Наличие миофибрилл, идущих вдоль волокна, обусловливает продольную исчерченность. Миофибриллы состоят из чередующихся светлых изотропных и темных анизотропных полос; светлые называются А-полосами, а темные - I-полосами. Эти полосы плотно уложены, что придает мышечному волокну вид поперечной исчерченности. Окончания волокон тупо закруглены.
Любая поперечно-полосатая мышца, например двуглавые мышцы, одета в покров из соединительной ткани, состоящий из трех компонентов. Эпимизий из соединительной ткани, содержащей большое количество жировых клеток, покрывает мышцу (влагалище мышцы) и, погружаясь, разделяет брюшко мышцы на пучки. Каждый пучок волокон в пределах мышцы окружен толстым слоем соединительной ткани - перимизием. В пределах пучка каждое поперечно-полосатое мышечное волокно окружено тонкой прослойкой, богатой кровеносными сосудами - эндомизием.
С помощью электронной микроскопии установлено, что волокна поперечно- полосатых мышц имеют сложное строение. Миофибриллы состоят из двух типов, смещающихся относительно друг друга, миофиламентов: тонких актин-содержащих и толстых миозин-содеращих. I-полосы состоят из тонких миофиламентов, а А-полосы - из обоих типов. Точно по центру каждой I-полосы расположена темная Z-линия, к каждой стороне которой прикрепляются актин-содержащие тонкие миофиламенты. Расстояние между каждой парой Z-линий называется саркомером, элементарная сократимая единица поперечно-полосатых мышц. Тонкие миофиламенты выступают в область А-полосы, причем один конец миофиламента свободен и находится между толстыми миофиламентами, что способствует возникновению светлой Н-зоны в середине А-полосы. Толстые миофиламенты тянутся через всю А-полосу и их концы свободны. Миофибриллы расположены в строго определенном порядке: каждый толстый миофиламент окружен шестью равноудаленными тонкими миофиламентами, которые расположены в форме шестиугольника.
При сокращении, длина миофиламентов не уменьшается, а увеличивается только лишь их степень смещения относительно друг друга. В результате этого, I-полоса уменьшается и Н-зона тоже становится очень узкой, однако длина А-полосы практически не изменяется. На поперечном срезе миофибрилла имеет 3 вида, в зависимости от уровня среза: только на уровне тонких миофиламентов, толстых миофиламентов или на уровне двух типов филаментов одновременно. Две эллиптические митохондрии окружают каждую миофибриллу на уровне I-полосы.
При исследовании под электронным микроскопом удается выяснить, что сарколемма состоит из плазматической мембраны и вместе с хорошо развитой базальной пластинкой и сетью тонких ретикулиновых волокон. Плазматическая мембрана вдавливается и проходит через волокно на уровне границы между А- и I-полосами в виде вытянутой трубочки (Т-трубочка). При сокращении волна деполяризации распространяется вдоль сарколеммы и благодаря Т-трубочкам более или менее синхронно достигает всех частей миофибриллы. Саркоплазматический гладкий эндоплазматический ретикулум опоясывает каждую миофибриллу несколько раз таким образом, что по бокам каждой Т-трубочки параллельно располагаются две терминальные цистерны. Эти структуры, видимые рядом в поперечном разрезе, называются триадой. Трубочки и цистерны обеспечивают изоляцию зоны катионов кальция, что происходит во время расслабления мышцы. Большинство мышц человека содержит три типа поперечно-полосатых волокон: красные, белые и промежуточные. Преобладающим типом являются красные волокна, которые характеризуются малой толщиной и обилием пигментированного белка - миоглобина, и относительно небольшим количеством миофибрилл. Красные, или слабо сокращающиеся волокна, выполняют повторяющиеся сократительные движения и характерны для мышц туловища, ответственных за поддержание позы. Белые волокна толще и содержат большее количество миофибрилл, но меньше миоглобина, чем красные волокна. Они способны сокращаться быстрее и сравнительно быстро устают, поэтому они больше подходят для коротких вспышек активности. Промежуточные волокна по размерам и характеристикам занимают промежуточное положение между красными и белыми волокнами.
Иннервация поперечно-полосатых мышц.
К каждому экстрафузальному поперечно-полосатому мышечному волокну подходит окончание аксона от двигательного альфа-мотонейрона в переднем роге спинного мозга и двигательных ядрах черепно-мозговых нервов. Некоторые альфа- мотонейроны иннервируют полдюжины поперечно-полосатых мышечных волокон, как в случае наружных глазных мышц, где необходим тонкий контроль. Другие альфа- мотонейроны могут иннервировать до 500 мышечных волокон в большой мышце. Один мотонейрон вместе с иннервированными им мышечными волокнами образует двигательную единицу. Нервно-мышечное (мионевральное) соединение или двигательная концевая пластинка обычно располагается в середине волокна, здесь оно утолщено и имеет локальное скопление ядер и митохондрий. Миелиновая оболочка аксона исчезает и он разветвляется на несколько оголенных грушевидных окончаний, которые погружены в углубления на поверхности мышечного волокна - это называется синаптической щелью. Здесь сарколемма очень складчата (соединительные складки или субнейральные щели). Лишь аморфный компонент сарколеммы (базальная пластинка) разделяет плазматические мембраны аксона и мышечного волокна и он содержит фермент ацетилхолинэстеразу. Окончание аксона содержит много митохондрий и скопления синаптических пузырьков, в которых находится нейромедиатор - ацетилхолин.
Мышечные веретена представляют собой инкапсулированные скопления истонченных мышечных волокон, лишь частично содержащих небольшое количество миофибрилл; данные волокна называются интрафузальными в отличие от экстрафузальных волокон мышц в большинстве случаев. Функционально, они являются проприоцептивными, действующими в качестве рецепторов растяжения и имеющими двойную иннервацию. Интрафузальные волокна иннервируются гамма-мотонейронами, которые имеют окончания в двигательных концевых пластинках. Данные волокна также имеют анулоспиральные сенсорные нервные, или первичные, окончания вокруг их центрального участка.
Ткань - это совокупность схожих по строению клеток, которые объединены общими функциями. Практически все многоклеточные организмы состоят из разных типов тканей.
Классификация
У животных и человека в организме присутствуют следующие типы тканей:
- эпителиальная;
- нервная;
- соединительная;
- мышечная.
Эти группы объединяют по несколько разновидностей. Так, бывает жировой, хрящевой, костной. Также сюда относятся кровь и лимфа. Эпителиальная ткань существует многослойная и однослойная, в зависимости от строения клеток можно выделить также плоский, кубический, цилиндрический эпителий и т. д. Нервная бывает только одного вида. А о мышечном типе ткани мы поговорим подробнее в этой статье.
Виды мышечной ткани
В организме всех животных выделяют три ее разновидности:
- поперечно-полосатые мышцы;
- сердечная мышечная ткань.
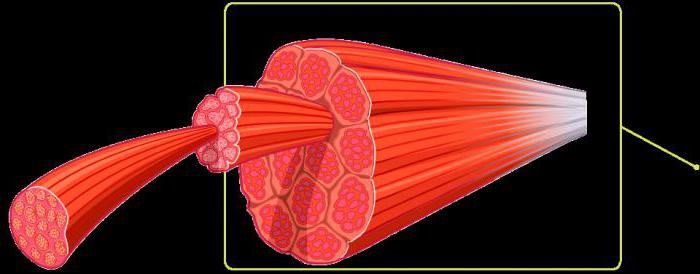
Еще одним общим признаком всех видов мышц является то, что они способны сокращаться, однако у разных видов этот процесс происходит индивидуально.
Особенности миоцитов
Клетки гладкой мышечной ткани, как и поперечно-полосатой и сердечной, обладают вытянутой формой. Кроме того, в них есть особые органоиды, которые называются миофибриллы, или миофиламенты. В них содержатся сократительные белки (актин, миозин). Они необходимы для того, чтобы обеспечить движение мышцы. Обязательным условием функционирования мускула, кроме наличия сократительных белков, также является присутствие в клетках ионов кальция. Поэтому недостаточное или избыточное употребление продуктов с высоким содержанием данного элемента может привести к некорректной работе мускулатуры - как гладкой, так и поперечно-полосатой.
Кроме того, в клетках присутствует еще один специфический белок - миоглобин. Он необходим для того, чтобы связываться с кислородом и запасать его.
Что касается органоидов, то кроме наличия миофибрилл особенным для мышечных тканей является содержание большого количества в клетке митохондрий - двумембранных органоидов, отвечающих за И это неудивительно, так как мышечному волокну для сокращения необходимо большое количество энергии, вырабатываемой при дыхании митохондриями.
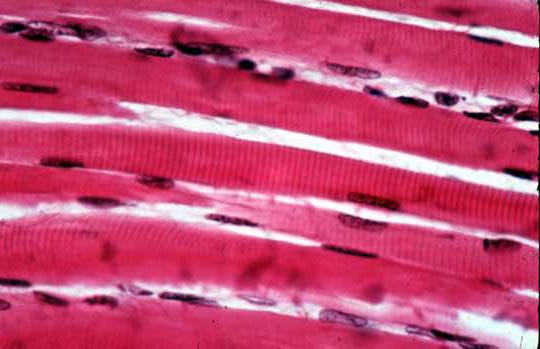
В некоторых миоцитах также присутствует более чем одно ядро. Это характерно для поперечно-полосатой мускулатуры, в клетках которой может содержаться около двадцати ядер, а иногда эта цифра доходит и до ста. Это связано с тем, что волокно поперечно-полосатой мышцы сформировано из нескольких клеток, объединенных впоследствии в одну.
Строение поперечно-полосатых мышц
Данный тип ткани еще называют скелетной мускулатурой. Волокна этого типа мышц длинные, собранные в пучки. Их клетки могут достигать нескольких сантиметров в длину (вплоть до 10-12). В них содержится много ядер, митохондрий и миофибрилл. Основная структурная единица каждой миофибриллы поперечно-полосатой ткани - саркомер. Он состоит из сократительного белка.
Главная особенность этой мускулатуры заключается в том, что она может контролироваться сознательно, в отличие от гладкой и сердечной.
Волокна данной ткани прикрепляются к костям с помощью сухожилий. Именно поэтому такие мышцы и называются скелетными.
Структура гладкой мышечной ткани
Гладкие мышцы выстилают некоторые внутренние органы, такие как кишечник, матка, мочевой пузырь, а также сосуды. Кроме того, из них формируются сфинктеры и связки.
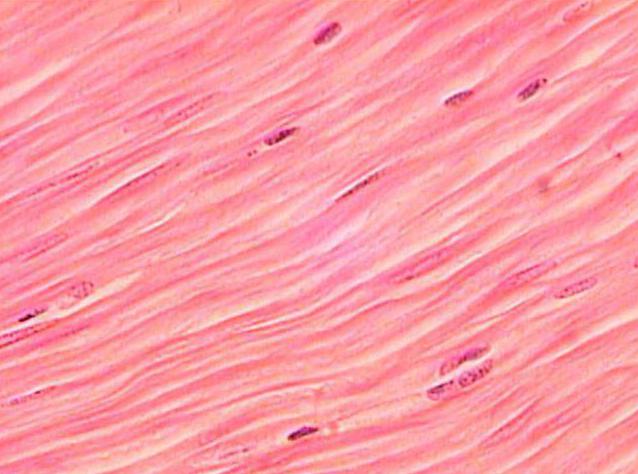
Гладкое не такое длинное, как поперечно-полосатое. Но толщина его больше, чем в случае со скелетными мускулами. Клетки гладкой мышечной ткани обладают веретоноподобной формой, а не нитевидной, как миоциты поперечно-полосатой.
Структуры, которые обесечивают сокращение гладких мышц, называются протофибриллами. В отличие от миофибрилл, они обладают более простой структурой. Но материал, из которого они построены, - все те же сократительные белки актин и миозин.
Митохондрий в миоцитах гладкой мускулатуры также меньше, чем в клетках поперечно-полосатой и сердечной. Кроме того, в них содержится только одно ядро.
Особенности сердечной мышцы
Некоторые исследователи определяют ее как подвид поперечно-полосатой мышечной ткани. Их волокна и вправду во многом похожи. Клетки сердца - кардиомиоциты - также содержат несколько ядер, миофибриллы и большое количество митохондрий. Данная ткань, как и способна сокращаться намного быстрее и сильнее, нежели гладкая мускулатура.
Однако основной особенностью, отличающей сердечную мышцу от поперечно-полосатой, является то, что она не может контролироваться сознательно. Сокращение ее происходит только автоматически, как и в случае с гладкими мышцами.
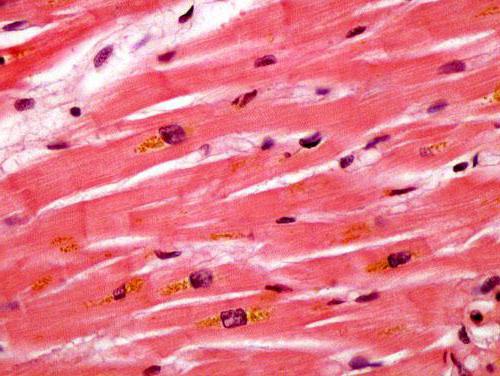
В составе сердечной ткани, кроме типичных клеток, присутствуют также секреторные кардиомиоциты. Они не содержат в себе миофибрилл и не сокращаются. Эти клетки отвесают за выработку гормона атриопептина, который необходим для регуляции артериального давления и контроля объема циркулирующей крови.
Функции поперечно-полосатых мышц
Основная их задача - перемещение тела в пространстве. Также это перемещение частей тела относительно друг друга.
Из других функций поперечно-полосатых мышц можно отметить поддержание позы, депо воды и солей. Кроме того, они выполняют защитную роль, что особенно касается мышц брюшного пресса, предотвращающих механическое повреждение внутренних органов.

К функциям поперечно-полосатой мускулатуры можно также причислить регуляцию температуры, так как при активном сокращении мышц происходит выделение значительного количества тепла. Вот почему при перемерзании мышцы начинают непроизвольно дрожать.
Функции гладкой мышечной ткани
Мускулатура данного вида выполняет эвакуаторную функцию. Она заключается в том, что гладкие мышцы кишечника проталкивают каловые массы к месту их выведения из организма. Также эта роль проявляется при родах, когда гладкие мышцы матки выталкивают плод из органа.
Функции гладкой мышечной ткани этим не ограничиваются. Также немаловажна их сфинктерная роль. Из ткани данного вида формируются специальные круговые мышцы, которые могут смыкаться и размыкаться. Сфинктеры присутствуют в мочевых путях, в кишечнике, между желудком и пищеводом, в желчном пузыре, в зрачке.
Еще одна важная роль, которую играют гладкие мышцы, - формирование связочного аппарата. Он необходим для поддержания правильного положения внутренних органов. При понижении тонуса этих мышц может происходить опущение некоторых органов.
На этом функции гладкой мышечной ткани заканчиваются.
Предназначение сердечной мышцы
Здесь, в принципе, особо говорить не о чем. Основная и единственная функция этой ткани - обеспечение циркуляции крови в организме.
Вывод: различия между тремя видами мышечной ткани
Для раскрытия этого вопроса представляем таблицу:
| Гладкая мускулатура | Поперечно-полосатые мышцы | Сердечная мышечная ткань |
| Сокращается автоматически | Может контролироваться сознательно | Сокращается автоматически |
| Клетки удлинненные, веретеноподобные | Клетки длинные, нитевидные | Удлинненные клетки |
| Волокна не объединяются в пучки | Волокна объединяются в пучки | Волокна объединяются в пучки |
| Одно ядро в клетке | Несколько ядер в клетке | Несколько ядер в клетке |
| Сравнительно небольшое количество митохондрий | Большое количество митохондрий | |
| Отсутствуют миофибриллы | Присутствуют миофибриллы | Есть миофибриллы |
| Клетки способны делиться | Волокна не могут делиться | Клетки не могут делиться |
| Сокращаются медленно, слабо, ритмично | Сокращаются быстро, сильно | Сокращаются быстро, сильно, ритмично |
| Выстилают внутренние органы (кишечник, матка, мочевой пузырь), формируют сфинктеры | Крепятся к скелету | Формируют сердце |
Вот и все основные характеристики поперечно-полосатой, гладкой и сердечной мышечных тканей. Теперь вы ознакомлены с их функциями, строением и главными различиями и сходствами.
Введение
Мышцы, органы тела животных и человека, за счет сокращения и расслабления которых происходят все движения тела и внутренних органов. Мышцы образуются мышечной тканью в сочетании с другими тканевыми структурами - соединительнотканными компонентами, нервами и кровеносными сосудами.
Мышечная ткань состоит из мышечных клеток, которым в наибольшей степени присуще свойство сократимости. Издавна эти клетки называют мышечными волокнами; данный термин указывает лишь на то, что мышечные клетки имеют вытянутую форму. Сократимость (одно из основных свойств животных клеток, которое наиболее развито в мышечных тканях) мышечных волокон обеспечивается аппаратом, образованным сократительными белками (актином и миозином), взаимодействие которых, протекающее с использованием энергии (АТФ), приводит к сокращению клеток (укорочению). Вслед за сокращением наступает расслабление, и тогда они возвращаются к своей исходной длине. Благодаря такому свойству клеток мышечной ткани достигается все многообразие двигательных функций организма и протекающих в нем механических процессов.
Существует три типа мышечной ткани и соответственно мышц, различающихся по структуре мышечных волокон и характеру иннервации:
1. Скелетная (поперечно-полосатая) мышечная ткань
2. Сердечная поперечно - полосатой мышечная ткань
3. Гладкая мышечная ткань
Скелетная (поперечно-полосатая) мышечная ткань -- упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов; один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания.
Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) диаметром от 50 до 100 мкм. Клетки многоядерные, содержат до 100 и более ядер. Микроскопическое изучение показало, что волокно скелетной мышцы по всей длине имеет регулярную поперечную исчерченность в виде чередующихся светлых и темных участков, (поперечнополосатая мышечная ткань образована мышечными клетками, содержащими миофибриллы, которые состоят из миозиновых и актиновых протофибрилл, взаимное положение которых создает поперечную исчерченность),что послужило основанием для другого названия - поперечнополосатые мышцы.
Функции скелетных мышц находятся под контролем центральной нервной системы, т.е. контролируются нашей волей, поэтому их называют также произвольными мышцами. Однако они могут находиться в состоянии частичного сокращения и независимо от нашего сознания; такое состояние называют тонусом. мышечный ткань волокно
Сердечная поперечно - полосатая мышечная ткань
Структурно-функциональной единицей сердечной поперечно - полосатой мышечной ткани является клетка - кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
Типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
Атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50-120 мкм в длину, шириной 15-20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть - функциональный синтиций. Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (дисков) между кардиомиоцитами нет. Вставочные диски - это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы - аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов - атипичные кардиомиоциты образуют проводящую систему сердца, состоящую из:
Синусо-предсердный узел;
Предсердно-желудочковый узел;
Предсердно-желудочковый пучок (пучок Гиса), ствол, правую и левую ножки;
Концевые разветвления ножек - волокна Пуркинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичных рядом особенностей:
Они крупнее (длина 100 мкм, толщина 50 мкм);
В цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
Плазмолемма не образует Т-канальцев;
Во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
· Р-клетки (пейсмекеры) - водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов - миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнее из целомического эпителия этих участков.









